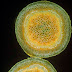
For hay fever sufferers, this can be one of the most miserable times of the year, thanks to grasses and trees that produce vast quantities of wind-borne pollen. Nevertheless, pollen grains are beautiful natural objects. The pollen grains above belong to a tropical plant called cup-and-saucer vine Cobaea scandens, which is pollinated by bats rather than by wind (do bats suffer from hay fever? Probably not...).
I stained the pollen with a fluorescent dye called acridine orange, which binds to the surface of the pollen grain and fluoresces yellow when you shine blue light on it - revealing this exquisite pattern of hexagons and pores (showing green here). Each pollen grain is about one fifth of a millimetre in diameter. Plant genera can often be identified by the distinctive pattern on their surface.
The outer surface of pollen is full of minute pits and chambers that contain proteins that allow a plant to recognise pollen of its own species when it lands on its stigma, and reject foreign pollen. It's these proteins that quite literally get up your nose, trigger an allergenic reaction and set you off sniffing and sneezing.
The outer casing (known as the exine) is made up of a polymer called sporopollenin, which is incredibly resistant to biodegradation - which is why palaeobotanists can recover ancient pollen samples from deep in peat bogs and lake beds and extract and identify pollen samples from plants that grew there tens of thousands of years ago. It's a branch of botany that has given some very useful insights into how plant species distributions have changed during periods of rapid climate change, like the one we are experiencing now: studying the past in this way gives an insight in what is likely to happen to plant species in the future.

Since the exine of pollen is so resilient, it passes through the gut of insects unharmed, although the pollen contents are digested. Yes, that little white speck on this bumblebee's tail is bee-poo, made up of empty exines of pollen that it has eaten. Many hoverflies feed almost exclusive on pollen, leaving little piles of hoverfly poo on leaves, and I know of at least one enterprising entomologist who has collected and analysed this, in order to study hoverfly's pollen diet.
During the Vietnam war Yellow Rain - yellow specks coating plants in the jungle - was believed to be the result of Communist chemical warfare. Subsequent anaysis showed that it was bee faeces, produced by vast swarms of bees that sometimes rose into the air and defecated in unison.
















.jpg)